Кора головного мозку є найбільш складною структурою людського мозку. Він має широкий спектр функцій, включаючи планування та ініціацію рухової активності, сприймання і усвідомлення сенсорної інформації, навчання, пам’ять, концептуальне мислення, усвідомлення емоцій і багато іншого. Виконання всіх цих функцій обумовлено унікальним багатошаровим розташуванням нейронів. Цитоархитектоника кори великих півкуль головного мозку – це їх клітинна організація.
Структура
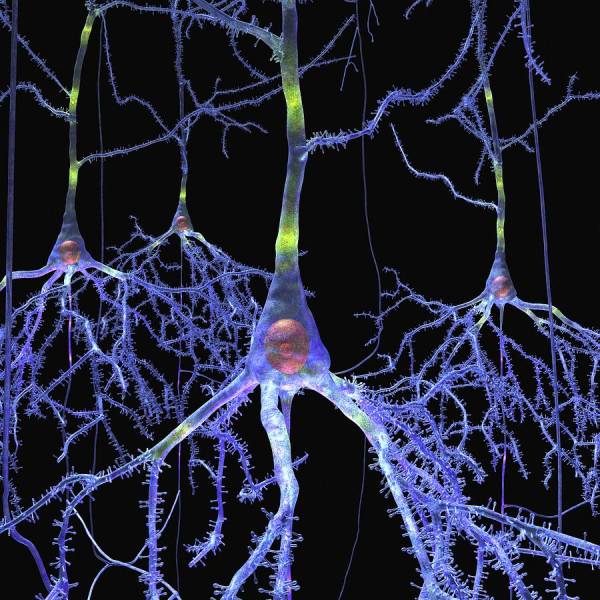
Кора головного мозку складається з сотень мільярдів нейронів, і всі вони являють собою різні варіації тільки трьох морфологічних форм: пірамідальні (пірамідні) клітини, веретеноподібні клітини і зірчасті (зернисті клітини). Інші типи клітин, видимих в корі, є модифікацією одного з цих трьох типів. Також там є горизонтальні клітини Кахаля-Ретциуса і клітини Мартинотти.
Пірамідальні клітини в цитоархитектонике кори півкуль становлять до 75 % клітинного компонента і є основними вихідними нейронами. Вони розрізняються за розміром від маленьких до гігантських. Вони зазвичай мають один апікальний дендрит, який йде до поверхні кори, і кілька базальних дендритів. Кількість останніх варіюється в широких межах, але, як правило, існує більше трьох-чотирьох первинних дендритів, що розгалужуються на наступні покоління (вторинних, третинних тощо) Зазвичай в них є один довгий аксон, який покидає кору і входить в подкорковое біла речовина.
Веретеноподібні клітини зазвичай розташовуються в самому глибокому кортикальному шарі в цитоархитектонике кори мозку. Їх дендрити виступають в напрямку поверхні кори, тоді як аксон може бути комиссуральным, асоціативним або орієнтованим на проекцію.
Зірчасті (зернисті) клітини зазвичай малі, і оскільки процеси проектуються у всіх площинах, вони нагадують зірку. Вони розташовані по всій корі, крім самого поверхневого шару. Процеси дуже короткі і проектуються локально в кору, можуть модулювати активність інших коркових нейронів. Виходячи з наявності дендритних шипів (невеликих цитоплазматичних випинань), деякі з них називають колючими клітинами. У їх дендритів є шипи, і вони в основному розташовуються в шарі IV, де вони виділяють глутамат, який є збудливим нейромедіатором, тому вони є функціонально збудливими интернейронами. Інший тип клітин виділяє гамма-аміномасляна кислота (ГАМК), яка є найбільш потужним інгібуючим нейротрансмітером центральної нервової системи, тому вони функціонують як інгібуючі интернейроны.
Горизонтальні клітини Кахаля-Ретциуса видно тільки в самої поверхневої частини кори. Вони дуже рідкісні, і тільки в невеликій кількості можуть бути знайдені у дорослому мозку. У них є один аксон і один дендрит, обидва вони синапсируются локально в поверхневому шарі.
Клітини Мартинотти є багатополюсним нейронами, які найбільш щільно розташованих у найглибшому шарі кори. Їх численні аксони і дендрити рухаються до поверхні.
